19. 5 세포분열단계 진입
*Precipitous (갑작스러운) mitotic CDK의 활성화가 체세포분열을 개시한다.

-M-CDK는 CDK subunit의 inhibitory 인산화로 인해 DNA 복제가 완성될때까지 불활성상태를 유지한다.
-S와 G2기에서 CDK 소단위체의 인산화는 M-CDK의 활성을 제한한다.
-M 사이클린의 농도는 점점 올라가는데 M-CDK의 농도는 G2후기부터 갑자기 올라간다.
-M-CDK의 활성화는 Wee1의 불활성화, 그리고 Cdc25의 활성화로 인해 일어난다.
-M-CDK는 빠르게 Wee1 kinase를 불활성화 시키고 Cdc42 phosphatase를 활성화시키는양성피드백 loop에 의해 스스로 활성화된다.
→Polo kinase family : 방추사 형성 / 염색체 분리
→Aurora kinase family: 방추사 형성 과정에서 중요한 역할
*Mitotic CDK는 nuclear envelope의 해체를 촉진한다.
라민 tetramer →MPF → 인산화된 라민 이량체 (dimers)
(핵막을 구성하는 라민을 쪼갠다)
i) NPC (nuclear pore complex)는 prophase에 M-CDK에 의해 인산화되어 soluble한, 막에 붙어있는 NPC complex로 분리된다.
ii) M-CDK가 INM(핵 내막)단백질을 인산화한다
→ INM 단백질이 nucelar lamina와 chromatin (염색질) 에 결합하는것을 억제한다
iii) 라민의 인산화 (by M-CDK)는 스스로의 depolymerization 유도 → 핵 lamina 해체
iv) Chromatin condensation (by M-CDK의 인산화)
→Chromatin과 핵막 사이 결합 억제
*M-CDK는 방추사(mitotic spindle) 형성을 촉진한다.
Centriole 복제 (G1-S 기, S기에 centriole 성숙)
-Centrosome disjunction (중심체 분리) : G1기
-amphitelic attachment : 각 염색체가 bi-oriented 된 방식으로 방추사에 연결된다
<잘못 연결된 경우>
i) Merotelic attachment : 하나의 kinetochore이 두 개의 다른 극에서 나온 방추사에 결합
ii) Syntelic attachment: 하나의 kinetochore이 동일한 극에서 나온 방추사에 결합
iii) Monotelic attachment : 하나의 방추사만 붙음
-단백질 인산화효소인 Aurora B 와 CPC (chromosomal passenger complex)로 알려진 억제인자들이 kinetochore이 tension 받는지 감시한다. (방추사 연결되면 장력받기 때문)
-Aurora B가 동원체 일부 성분을 인산화 → microtubule과 결합능력 잃음.
동원체는 tension없을 때 Aurora B에 접근
kinetochoe에 장력이 걸리지 않으면 Aurora B는 kinetochore의 미세소관 결합 부위를 인산화시켜서 미세소관의 결합 친화도를 낮춘다.
*염색체 응축이 염색체 분리를 촉진한다
-Sister chromatid resolution: 엉친 자매 염색분체를 푸는 과정
-Condensin (콘덴신) → 염색체 응축시키는 역할. M-CDK에 의해 활성화된다.
19.6 체세포분열 완성: 염색체 분리, 분열기 끝내기
*Separase (세퍼레이즈)에 의해 중재되는 코헤신 cleavage는 염색체 분리를 개시한다.
-Separase (protease) : 코헤신 소단위체를 자른다 (Scc1와 Rad21)
*APC/C는 securin (시큐린)의 유비퀴틴화를 통해 활성화된다.
-후기(anaphase)로 넘어가기 전에 시큐린이 세퍼레이즈에 붙어서 세퍼레이즈를 억제한다.
-APC/CCdc20 는 mitotic CDK가 APC/C의 subunit들을 인산화시킴으로서 후기에 인산화된다.(그러나 모든 염색체가 방추사와 bi-oriented될때까지 인산화된 APC/C는 인산화되지 않는다.)
*M-CDK 불활성화는 분열기의 끝을 알린다!
*핵막 재형성
→ ER은 확장해 탈condensation하는 각 염색체와 결합하고 서로 융합되어 염색체 주위에 이중막을 형성한다. 탈인산화된 핵공의 subcomplex는 다ㅣ 핵공으로 reassemble 된다 → karyomere이라고 불리는 individual mini-nuclei를 만든다. 모든 karyomere의 핵막은 각 spindle pole과 융합하여 한 세트의 염색체를 포함하는 하나의 핵을 형성한다.
*Cytokinesis (세포질 분열)
-Spindle midzone에서 cell cortex로 보내진 신호가 세포질 분열이 일어날 site를 조절한다.
-Spindle의 microtubule이 spindle pole position에 대해 cleavage furrow에 위치한 cell cortex와 상호작용한다.
-세포질 분열의 개시 신호: M-CDK의 불활성화
19.7 세포 주기 조절의 Surveillance Mechanism
*Sensor 는 특정 세포주기의 과정에서 나타나는 결함들을 감지하고 그에 따라 신호 전달 경로를 활성화시킨다.
*Effector는 위의 sensor가 보낸 신호 전달 경로에 의해서 활성화 되고 결함을 고치는 과정을 개시시킨다. 또 결함이 고쳐지기 전까지 세포 주기를 멈춘다. 고칠 수 없는 결함이라면 세포자살 (apoptosis)를 진행한다.
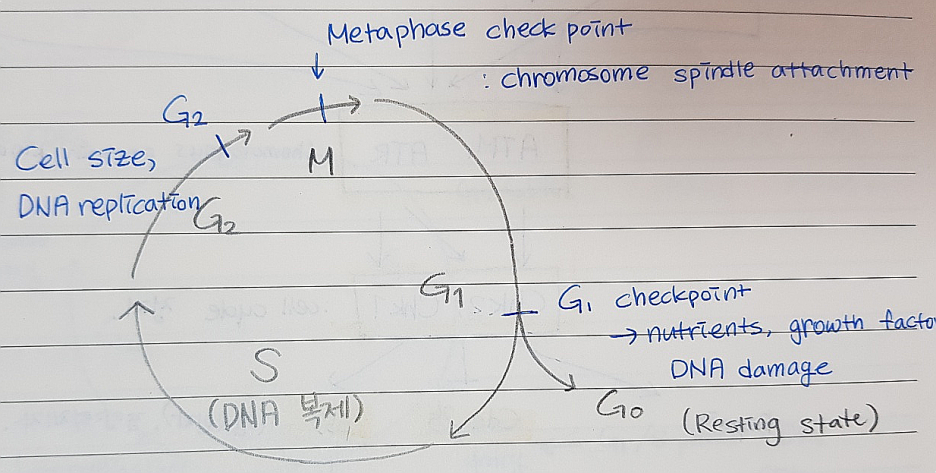
*Cdc13: 텔로미어 복제에 쓰인다. (온도에 민감함)
cdc13만 변이되면 restrictive 온도에서 cell arrest (세포주기 더 진행 못하게 막는것) 일어남, 하지만 double mutant (cdc13, RAD9)는 arrest되지 않는다.
*RAD9: viability에는 중요하지 않지만 결실되면 DNA damaging agent에 민감해진다.
→ 체크포인트 경로 (checkpoint pathway) : RAD9 결실되면 세포주기 정지 신호가 제대로 전달되지 않는다.
*Cln3: G1기 사이클린
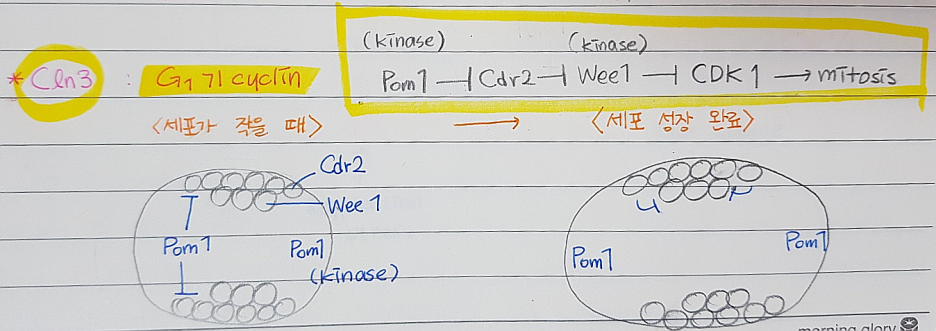
*DNA 에 손상을 입었을때
: I) 수선 시스템 (repair system)
II) 세포자살 경로 / programmed cell death
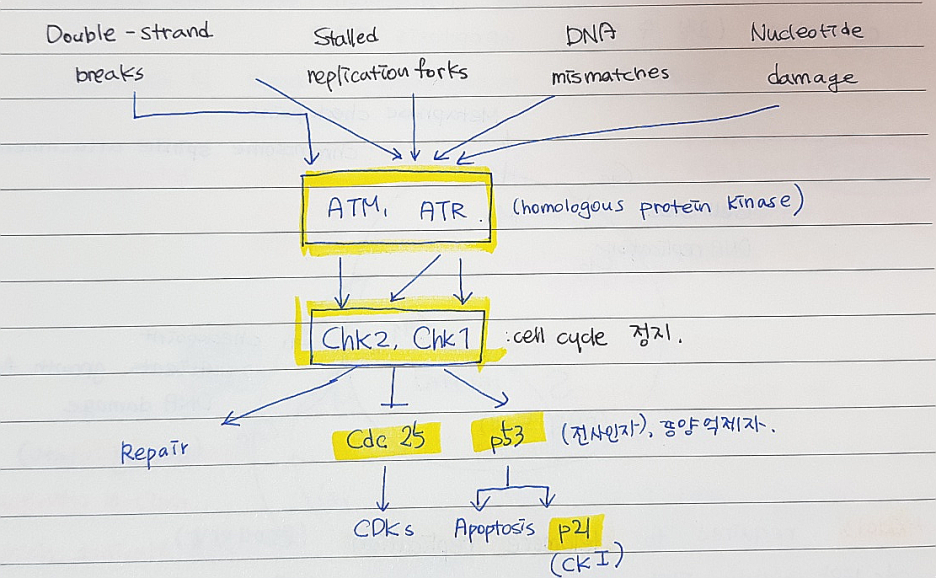
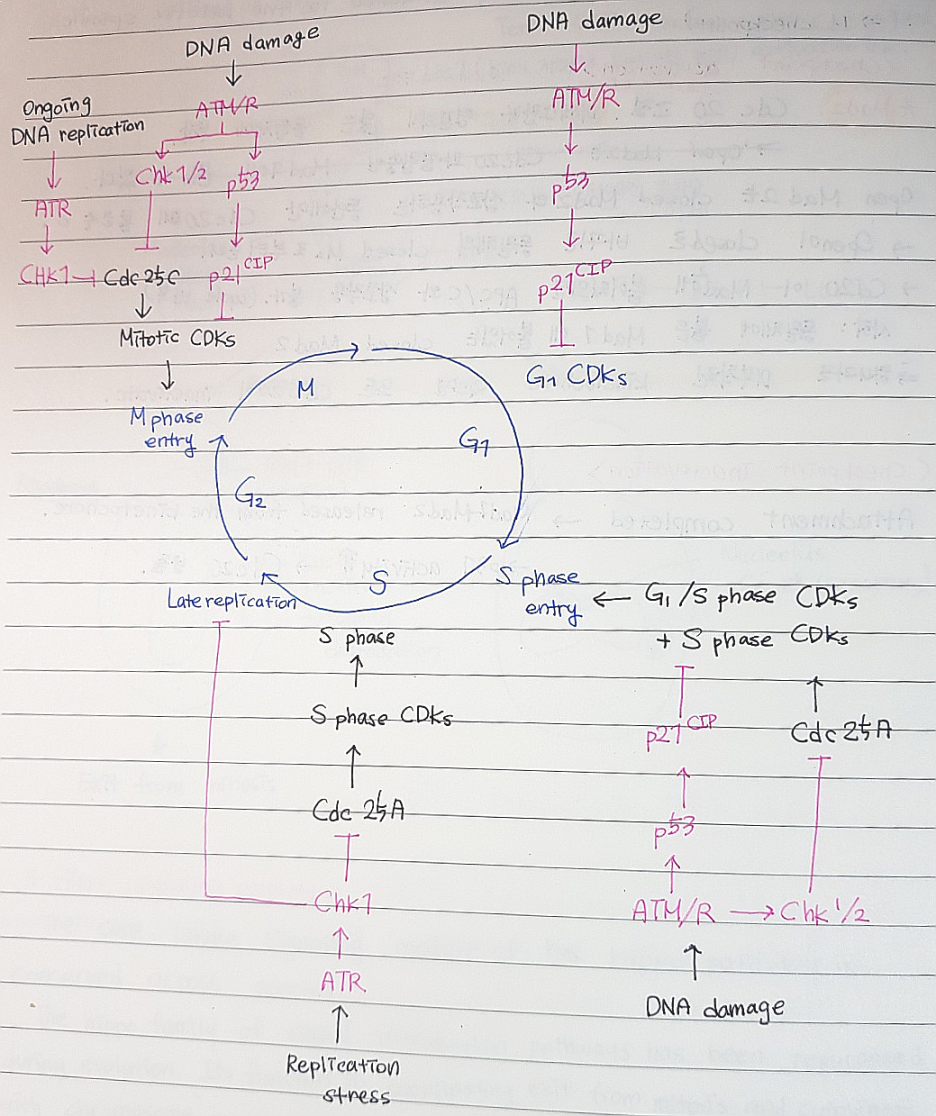
<DNA 손상 체크포인트 조절의 개요>
*Spindle Assembly Checkpoint = M 체크포인트
<체크포인트 활성화>
*Mad2: Cdc20 (세큐린 떼는 애) 조절. 미세소관과 연결되지 않은 동원체에 부착한다
Open Mad2는 closed Mad2와 상호작용하는동안에만 Cdc20에 붙을 수 있다
→ open이 closed로 바뀌고 동원체의 closed M2로부터 분리
→ Cd20이 Mad2에 붙어있으면 APC/C와 상호작용 불가 (Cycle 반복)
시작: 동원체에 붙은 Mad1에 붙어있는 closed Mad2
→하나라도 미부착된 kinetochore 있으면 모든 cdc20이 불활성화된다.
<체크포인트 불활성화>
attachment 완료 → Mad1-Mad2가 kinetochore로부터 분리 → p31 활성도 높아짐 → Cdc20 방출
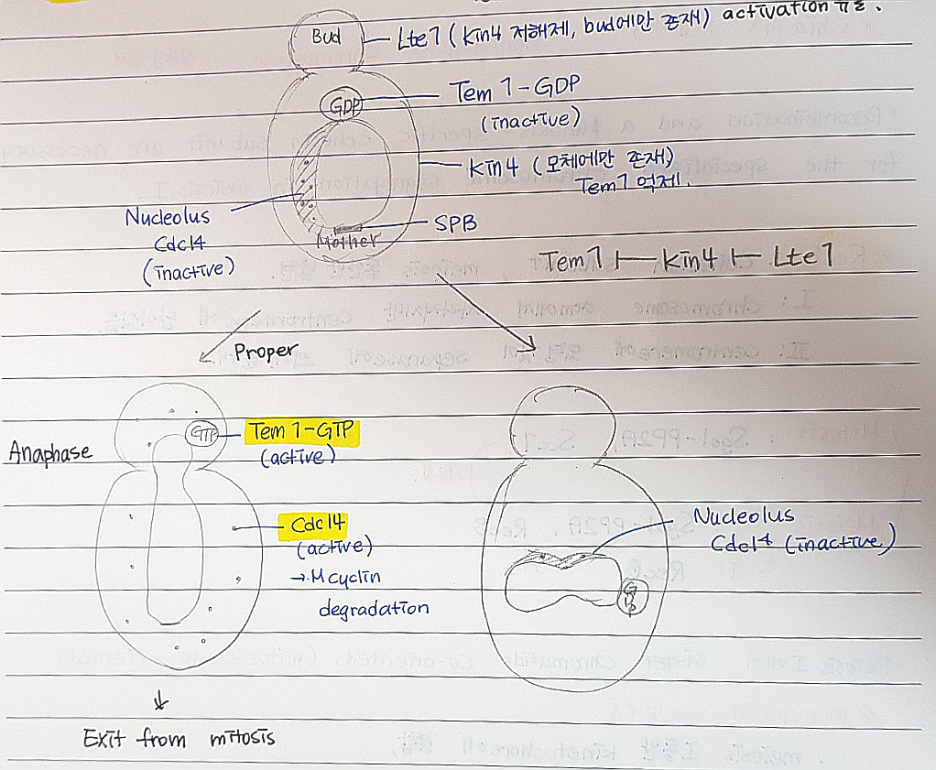
*Spindle position Checkpoint pathway : 핵이 정확히 이등분 되도록
-Tem1: GTPase에 속함. mitotic exit network조절.
Tem1-GTP: nucleolus 에서 세포질로 Cdc14 해리, 활성화 유도.
*Hippo signaling pathway
-이 Hippo 신호전달경로의 중심이 되는 인산화효소는 종 간에 보존되었다.
-The hippo family of signal transduction pathways has been repurposed during evolution. Its function in coordinating exit from mitosis and cytokinesis with chromosome segregation in fungi has been replaced with coordination of tissue growth with tissue organization in metazoans.
19. 8 Meiosis (감수분열): 특별한 세포분열
이 소단원에서는 그냥 mitosis와 meiosis의 차이점을 간단하게 다뤘다.
-chiasma: 교차점
-homologous chromosome : 상동염색체
*감수분열과 재조합 : 제1 감수분열에서의 염색체 분리를 위해서는 특별한 코헤신 subunit이 필요하다.
-Rec8: 코헤신 subunit, meiosis 동안만 발현.
I: chromosome arm에서 사라지지만 centromere에 남아있음.
II: centromere에 있던 것이 separase에 의해 분해.

*감수1분열에서 자매염색분체: co-oriented (체세포분열에서는 bi-oriented였음)
-Monopolin complex: 감수1분열동안 kinetochore에 결합
*감수 1분열과 2분열 사이에 DNA 복제는 억제된다.
: 1기 후기에서 meioric CDK의 활성도가 여전히 높음. (불완전한 CDK의 불활성화).→ MCM 헬리케이스 로딩 불가
'공부하자 > 세포생물학-Lodish' 카테고리의 다른 글
| [세포생물학] Chapter 19. Eukaryotic Cell Cycle 1~4 정리 (2) | 2024.11.20 |
|---|---|
| Chapter 18. Microtubules and Intermediate filaments 7~8 정리 (7) | 2024.11.12 |
| [세포생물학] Chapter 18. Microtubules and Intermediate filaments 6정리 (0) | 2024.11.07 |
| [세포생물학] Chapter 18. Microtubules and Intermediate filaments 4~5 정리 (0) | 2024.10.29 |
| [세포생물학] Chapter 18. Microtubules and Intermediate filaments 1~3 정리 (1) | 2024.10.22 |



