17.5 Myosins: Actin-based Motor proteins (운동 단백질!)
들어가기 전에) Sacomere (근절) < myofibril < Muscle fiber < muscle
점점 오른쪽으로 갈 수록 모여서 만들어진다 (큰 단위)
Multiple myosin molecules are oriented in opposite directions.
chain들은 3개의 유전자로 암호화 되어있다.

위의 사진은 미오신 II의 모습이다. 미오신 II는 ATPase 활성이 있다. 위에 파란색과 분홍색으로 표시한 건 자르는 물질과 잘려지는 단위체를 표시한 것이다. 키모트립신을 처리하면 HMM(점선 왼쪽)과 LMM(점선 오른쪽)으로 나뉘어지고, Papain을 처리하게 되면 S1, S2로 나뉘어진다. (HMM: heavy meromyosin)
Light chain (경쇄)는 regulatory 와 essential 두 부분으로 구분되어 있고, 미오신II 은 각각 두 개씩 가지고 있다. 헤드 부분에는 액틴 결합부위가 있어서 액틴이 결합할 수 있다. 액틴 결합부위 (actin binding site)에 ATPase 활성이 있기 때문에 이 부위가 중요하다.
대부분의 미오신 head는 (+)말단 쪽을 향해서 걸어간다.
*미오신의 종류
-Myosin I
: 미오신 II를 반 자른 것 처럼 생겼다. (하나의 헤드만 가짐.) membrane association, endocytosis에 관여한다.
associate directly with membranes through lipid interactions. (-)에서 (+)로 이동한다.
-Myosin II
:위에서 나온 것처럼 생겼다. 보이는 것처럼 헤드가 두 개 있어서 bipolar filament이다.
Contraction에 쓰이며, 미오신 II의 neck 길이가 움직이는 속도를 결정한다. (길수록 빠름) processive.
-Myosin V
:Organelle transport에 쓰인다. 몸 속의 vesicle을 옮길 수 있다는 뜻이다. 또, 효모의 budding에서 쓰인다.
미오신 5번은 filament로 합성되지 않는다. 미오신V는 미오신 II랑 비슷하게 생겼는데, lightchain이 6개라서 미오신 II보다 훨씬 길다. 중쇄 (Heavy chain) 끝에는 domain이 있어서 vesicle 같은 옮길 물질들을 부착할 수 있다.
-Myosin VI
:미오신6번이 특이한 점은 얘만 이동방향이 반대라는 것이다. 얘만 (+)에서 (-)로 이동한다.
그래서 Endocytosis할 때 cell center를 향해 물질을 옮길 때 사용한다.
미오신은 어떻게 움직일까?
: ATP 가수분해에 의해 방출된 에너지를 이용해 액틴 필라멘트를 따라 움직인다.
i) ATP가 붙으면 미오신 헤드가 액틴 필라멘트에서 떨어진다.
ii) ATP → ADP + Pi (미오신 헤드가 돌아간다) → "cocked state"
iii) 액틴에 결합한다
iv) "Power stroke"가 일어난다 : ADP + Pi → ADP (액틴 움직임)
v) ADP released, 새 ATP 올 때까지 부착해있는다. (새로운 ATP 없으면 못떨어지고 계속 붙은 채로 있는다.)
17.6 Myosin-powered movements
tail 부분 → 어떤 물질을 수송할지 결정한다.
*Myofibril (근원섬유)
sacomere (근절)
-Thick filament : myosin II, bipolar filament 미오신 다발이 액틴보다 더 두껍다.
-Thin filament : 액틴 + binding protein.
I band: 미오신 없는 부분, A band: 미오신 있는 부분.(미오신의 길이)
+) Hypertrophic cardiomyopathy : heart wall muscle thickening.
heart contractile machinery 형성하는 단백질에 변이가 생긴다. 결과적으로 미오신 중쇄에 결함이 생김.
-Nebulin: 액틴에 붙어있는 단백질. thin filament의 길이를 결정한다.
-Titin: 미오신, head가 Z disk에 부착, thick filament를 중앙에 배열한다.
-Cap z: (+) capping (thin filament에서)
-Tropomodulin: (-) capping
이 네 가지 단백질들은 accessory protein들이다.
*Contraction 조절하기: Ca2+, Actin binding protein
-Sarcoplasmic reticulum (SR): 근육 세포의 칼슘이온 저장고라고 보면 된다. 소포체처럼 칼슘이온의 농도가 높다.
<Thin filament regulation> : 이건 skeletal muscle에서 일어난다고 생각하면 된다. (근육 수축을 위해)
Sarcolemma(근육세포의 plasma membrane)에 활동전위 발생 → Transverse tubule (Myofibril 주변 위치) → SR의 voltage-gated(전압의존성) 칼슘 채널 열림 → 세포질의 칼슘농도 상승 → 트로포미오신, 트로포닌의 변화 → 미오신-액틴 interaction → Contraction (근육수축)
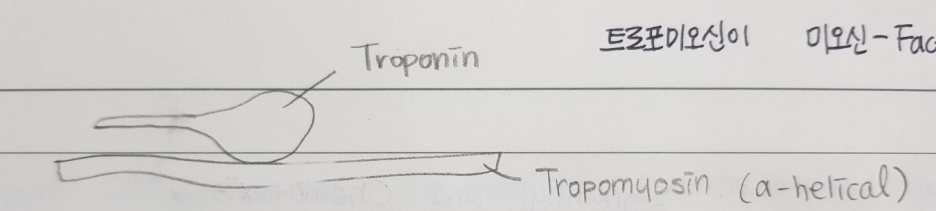
-Tropomyosin(트로포미오신): 약 40nm로 액틴 필라멘트의 약 7개 액틴 subunit과 결합한다.
-Troponin : TN-T, TN-I, TN-C 등이 있으며, TN-C는 Ca2+ binding subunit으로 트로포미오신의 위치를 조절한다. 칼슘이온이 결합하면 액틴의 myosin binding site가 노출된다.
*Contractile bundle in nonmuscle cells: actin + myosinII
Skeletal muscle이 아닌 경우에는 어떨까?
i) Adherence belt (=circumferential belt)
ii) Stress fiber : cell adhesion에 중요하다. 말단: focal adhesion (인테그린 포함)
iii) Contractile ring (수축환) : actin filament. Myosin II is concentrated on the contractile ring while myosin I is localized at the poles.
미오신에 의존한 매커니즘들이 smooth muscle과 nonmuscle 세포에서 수축을 조절한다.
→ 미오신 regulatory light chain phospho/dephosphorylation
<Thick filament regulation>
Smooth muscle myosin II (여기서 i, ii의 두 가지 경로로 진행할 수 있다.)
i) → RLC 인산화 →(칼슘농도로 조절되는 미오신 LC kinase)→ bipolar filament →수축
ii)→ dephosphorylation → folded → ATP cycle 불활성
수축은 여기서 더 느리다!
Skeletal muscle은 nerve impulse로만 수축하지만, smooth/nonmuscle 은 다른 기작으로 더 조절 될 수 있다 (Rho kinase 처럼)
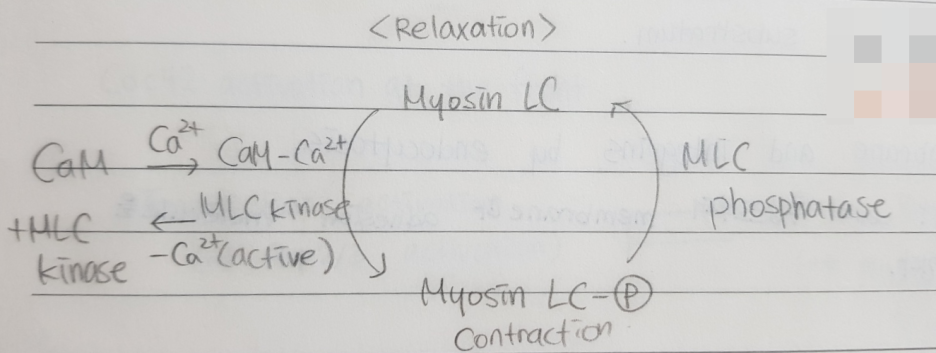
요약:
Skeletal muscle : Ca2+, 트로포미오신-트로포닌 복합체에 의해 조절된다.
Smooth muscle: 미오신 II의 on/off 상태 cycling으로 조절된다.
17.7 Cell migration: Mechanism, signaling, and chemotaxis
전체적인 과정은 :
Extension → 부착 →translocation → De-adhesion, endocycling recycling.
i) Membrane extension: Leading edge (Listeria같은 매커니즘으로 움직인다)
: 액틴 중합이 일어난다. 앞서 배운 Arp 2/3 complex에서 nucleation을 한 뒤 cofilin, profilin을 이용해 액틴의 polymerization을 조절한다. 발을 뻗는 작용이라고 생각하면 됨.
ii) Cell-substratum adhesion
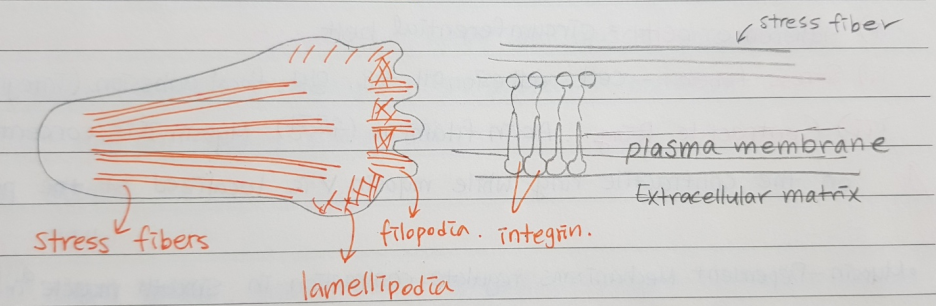
Leading edge의 액틴 다발이 focal adhesion에 연결된다.
Cell-adhesion molecule : integrin (막단백질) 인테그린을 통해서 세포막에 붙는다.
iii) Cell-body translocation
미오신 II를 통해 translocation 한다. 이동하기 위해 다리를 옮기는 행동이라고 생각하면 된다.
cell cortex의 후방에서 이루어진다.
iv) Breaking cell attachment
Focal adhesion이 떨어진다. 인테그린은 재사용되고, 꼬리 부분이 앞으로 이동한다.(할 일이 끝나서?)
기계적 force(by cytoskeleton)와 cell adhesion에서 생긴 Resisting force가 이 움직임을 조절한다.
→ Cell locomotion results from traction forces exerted by the cell on the underlying substratum.
v) Recycling of membrane and integrins by endocytosis
Endocytic cycle: 세포의 후방으로부터 막과 adhesion molecule을 가져가 앞에 삽입시킨다. (말그대로 재료 재활용 하는것임)
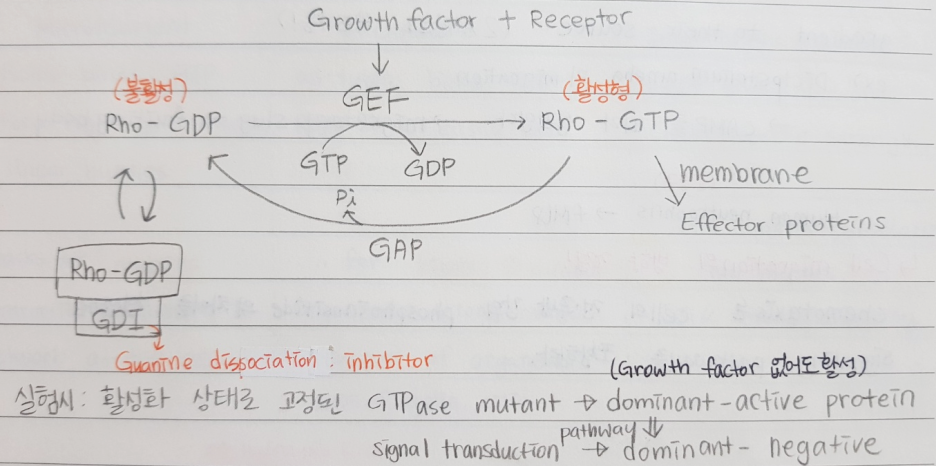
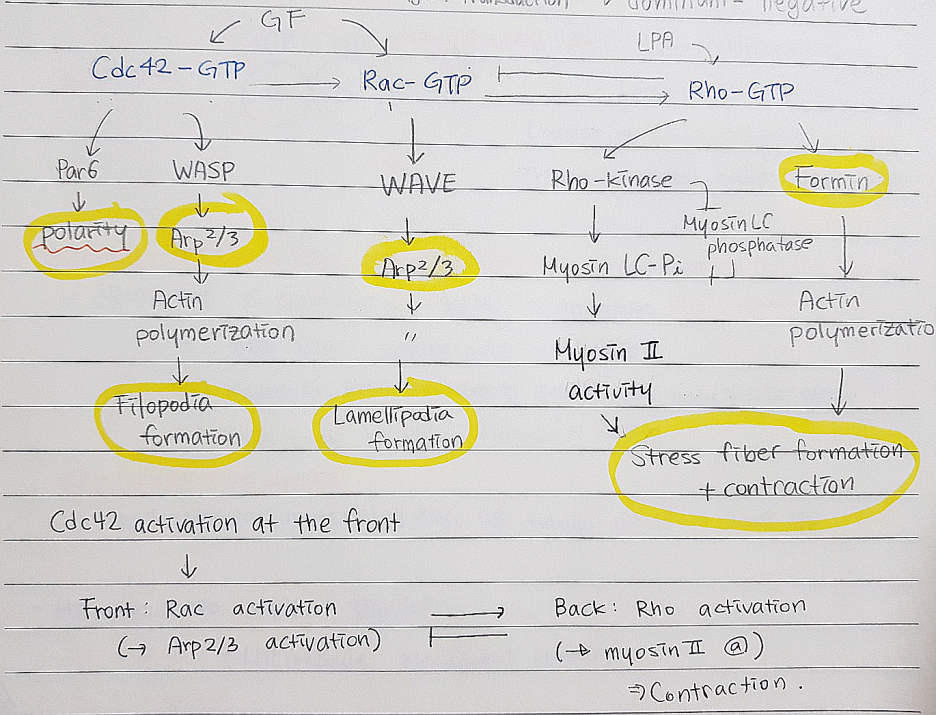
*Small GTPase of the Rho family: Cdc42, Rac, Rho
→얘네는 GTP 붙었을 때가 활성형이다.
*Chemotaxis (주화성)
세포들은 수용성 작은 물질들을 sense하고 그 농도 기울기에 따라 이동할 수 있다.(source 쪽으로)
단 2%농도 만으로 주화성을 유도할 수 있다!
예시) Dictostelium 아메바의 migration
이 경우에는 cAMP의 농도기울기를 따라 이동한다. → migratory slug →fruiting body
예시2) 사람의 호중구는 fMLP 농도를 따라 이동한다.
우리가 배우고 있는 CELL Migration에서는 주화성이 migration의 방향을 조절하는 데 쓰인다.
이 주화성은 세포의 전, 후방 간의 phosphoinositide의 농도차를 형성하는 신호 pathway를 포함한다.



